Flagelo

O flagelo é um motor molecular (nanomáquina) utilizado por organismos microscópicos, tais como bactérias e protozoários, para propulsionar-se através de um meio aquoso. O flagelo bacteriano é um filamento helicoidal que gira como um motor de popa, enquanto o flagelo eucariota é uma estrutura em forma de chicote que chicoteia para frente e para trás.
Máquinas moleculares, tais como o flagelo servem como exemplos poderosos de tipos de design inteligente sofisticado que podem ser encontrados mesmo no mais simples dos organismos. Defensores do DI, como Michael Behe têm ajudado a elucidar o flagelo, como prova irrefutável de DI, ilustrando a função irredutivelmente complexa de seus sistemas.
Os físicos modernos olham para motores moleculares, como o flagelo bacteriano, em uma tentativa de melhorar os métodos de concepção de motores modernos, na esperança de alcançar a eficiência funcional evidente dentro da criação.
| “ | Segundo o Instituto Americano de Física: Os projetos estruturais e os mecanismos funcionais revelados na maquinaria complexa do flagelo bacteriano poderiam fornecer novas tecnologias que viriam a se tornar uma base para a futura nanotecnologia, do qual deveremos ser capazes de encontrar muitas aplicações úteis.[1] | ” |
Flagelo bacteriano
O flagelo bacteriano é composto por cerca de 25 proteínas diferentes, cada um delas em várias cópias, variando desde umas poucas a algumas dezenas de milhares. É accionado por um motor rotativo com um diâmetro de apenas 30 a 40 nm. Muito parecido com o moderno motor rotativo, o flagelo bacteriano é composto por várias partes diferentes que realizam diferentes funções, tais como um motor rotativo, um bucha, um eixo de transmissão, um regulador de rotação por switch, uma junta universal, uma hélice helicoidal e um promotor rotativo para auto-montagem.[1] O filamento do flagelo bacteriano é um tubo oco composto de proteína, e é de apenas 20 nanômetros de espessura. É helicoidal, e tem uma curva acentuada do lado de fora da membrana externa chamado "gancho" que permite que a hélice possa apontar na direcção oposta da célula. Um veio corre entre o gancho ea estrutura basal, passando através de anéis de proteínas em membranas da célula que actuam como rolamentos.[Carece de fontes]

O motor pode funcionar a 6.000 a 17.000 rpm, mas com um filamento ligado normalmente só atinge 200 a 1000 rpm.[Carece de fontes]. O motor acciona a rotação do flagelo em cerca de 300 Hz, a um nível de potência de 10-16 W com eficiência de conversão de energia perto de 100 %.[1]
Origem
Kenneth Miller, um biólogo evolucionista bem conhecido da Universidade Brown, aponta que determinados subsistemas do flagelo bacteriano ainda estariam em ordem de funcionamento se outras peças foram removidas. O sistema de mobilidade geral flagelar requer cerca de 50 tipos diferentes de proteínas (e genes subjacentes ao código para elas). No entanto, é bastante interessante notar que 10 destes genes e a estrutura resultante dentro do sistema de motilidade flagelar codificam também para o que é conhecido como um sistema secretor tipo III (TTSS). O TTSS é usado como uma toxina injetora por algumas bactérias especialmente desagradáveis que atacam tanto os animais quanto as plantas. Portanto, Kenneth Miller afirma que é um erro usar o sistema flagelar como um exemplo de uma máquina verdadeiramente de complexidade irredutível uma vez que cerca de 40 partes diferentes poderiam ser removidas da máquina sem uma perda completa de função. Miller também aponta que a maioria das partes de proteína do sistema flagelar têm outras funções como partes de outros sistemas dentro das bactérias.[2]
E agora? Miller refutou a noção de Behe de complexidade irredutível? Com certeza parece que sim - à primeira vista de qualquer maneira. No entanto, o que Miller aparentemente deixa de considerar é que a função da motilidade flagelar ainda é irredutivelmente complexa, independentemente de outras funções de subsistemas serem ou não serem mantidas com a redução de vários sistemas flagelares. Sem um número considerável de peças de proteínas especificamente dispostas a função de motilidade flagelar não pode existir. Na verdade, todos os sistemas da função são irredutivelmente complexos. Não importa se a função do subsistema é mantida. Isto é como argumentar que a função de motilidade de um automóvel não é irredutivelmente complexa porque as luzes continuam a trabalhar mesmo se o motor ou os pneus ou o eixo de acionamento são removidos.
Mas, que tal noção de Miller que os subsistemas de trabalho podem ser facilmente adicionados para formar funções com mais e maiores requisitos mínimos de limiar estruturais? Esta noção é de fato verdade até certo ponto. Há muitos exemplos de evolução de baixo nível em ação. O próprio Miller aponta alguns destes em seu livro, Finding Darwin's God, de modo a incluir um experimento por Barry Hall. O experimento de Hall é de fato muito interessante. O que Hall fez foi eliminar os genes de um tipo de bactéria chamada E. coli que produziam uma proteína chamada de enzima lactase. Sem esta enzima lactase as bactérias não podiam mais digerir a lactose do açúcar para obter energia. Hall se perguntou se estas bactérias, se crescessem em um ambiente rico em lactose, evoluiriam a função da lactase novamente usando algum outro aspecto do seu pool genético. E, eles fizeram! Em apenas algumas gerações (provavelmente uma única geração) a colônia de alguns bilhões de bactérias evoluiu de uma completamente nova enzima lactase, com apenas um único ponto de mutação de outro gene. Este produto do gene não tinha a função de lactase antes. Assim, esta parece ser uma evolução real de uma nova função em tempo observável.
É claro, este é o lugar onde as descrições de Miller dos experimentos de Hall terminam. No entanto, o que Hall fez a seguir é o mais interessante. Hall imaginou o que aconteceria se ele apagasse o gene da lactase recém-evoluído. Será que as bactérias evoluiríam ainda outra nova enzima lactase? Hall realizou esse experimento e observou as bactérias por mais de 40.000 gerações - e elas não evoluíram outra enzima lactase, apesar de uma taxa de mutação muito elevada em uma população grande e em um ambiente altamente seletivo. Frustrado, Hall se referiu a esta colônia como tendo "limitado potencial evolutivo".[3]
Agora, o que é que limita o potencial evolutivo das bactérias de Hall? Como é que uma função relativamente simples não pôde ser encontrada por mutação aleatória e seleção baseada em função diversas vezes em tal colônia grande vivendo em um ambiente altamente seletivo?
A resposta, é encontrada na expansão das lacunas não-benéficas entre as sequências genéticas potencialmente benéficas quando se considera funções com maiores e maiores mínimos requisitos de limite estrutural. Como se vê, a evolução prossegue com bastante facilidade e muito rapidamente quando se trata de sistemas funcionais que exigem apenas poucas exigências de limite estrutural ou perda de um sistema pré-existente de interação (a maioria das formas de resistência aos antibióticos). Ocasionalmente mecanismos evolutivos produzem funções de alto nível em que algumas centenas de resíduos frouxamente especificados de aminoácidos são necessários (lactase, nylonase, etc.) No entanto, não existem exemplos de evolução observáveis em acção produzindo qualquer novo sistema de função que requeira mais de 1.000 resíduos de aminoácidos especificamente arranjados que trabalhem em conjunto, ao mesmo tempo. Não há um só exemplo de evolução para além deste nível em toda a literatura científica - nem um exemplo.[4]
Agora, considere que o sistema de motilidade flagelar requer cerca de 10.000 resíduos de aminoácidos especificamente dispostos de maneira justa, todos trabalhando juntos, ao mesmo tempo. O mais próximo "degrau" de função de subsistema benéfico não é nem remotamente perto dste nível de complexidade funcional. Mesmo se dois subsistemas pudessem ser colocados juntos para fazer o sistema flagelar, as chances de que fossem necessárias apenas algumas mudanças de resíduos são extremamente baixas. Mais provavelmente muitas dezenas de alterações de resíduos seriam necessárias. As chances de se ter as mudanças necessárias de aminoácidos de resíduos presentes em uma bactéria dentro de uma imensa população ao mesmo tempo são extremamente remotas da ordem de trilhões de trilhões de anos de tempo. Essas lacunas podem não parecer muito à primeira vista, mas quando consideradas mais de perto, as chances de serem alcançadas no que alguém iria considerar uma quantidade razoável de tempo são praticamente impossíveis.
A esta luz, consideram que o sistema TTSS é agora pensado como tendo evoluído a partir do flagelo totalmente formado - e não o contrário. Em 2000, Nguyen et. al. publicaram um artigo no Journal of Molecular Microbiology and Biotechnology intitulado, Phylogenetic analyses of the constituents of Type III protein secretion systems[5] Neste trabalho, os autores argumentam fortemente que o sistema TTSS evoluiu do flagelo. Claro, aqueles como Miller deixam de apontar tais conclusões publicadas na corrente principal da literatura científica. Mas tais conclusões deveriam ter sido óbvias desde o início. O que veio primeiro? - - um peixe das cavernas sem olhos ou um peixe em uma lagoa com os olhos? É claro que é muito mais fácil perder algo que já estava lá do que criá-lo em primeiro lugar. Lembre-se de Humpty Dumpty e todos os homens reis?
| “ | No livro A Origem das Espécies Charles Darwin afirmou: "Se pudesse ser demonstrado que existiu algum órgão complexo que não poderia ter sido formado por numerosas, sucessivas e ligeiras modificações, minha teoria seria totalmente invalidada." | ” |
Galeria

Micrografia eletrônica do H. pylori possuindo múltiplos flagelos.
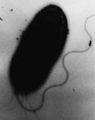
Uma bactéria com um flagelo em forma de chicote.
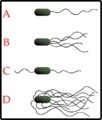
Exemplos de esquemas de arranjo de flagelos bacterianos. A-Monotrico; B-Lofhotrico; C-Anfitrico; D-Peritrico.







Referências
- ↑ 1,0 1,1 1,2 Self-Assembly of Bacterial Flagella by the American Institute of Physics
- ↑ Dr. Sean D. Pitman (Maio, 2006). The Evolution of the Flagellum. Página visitada em 7 de Julho de 2012.
- ↑ Sean D. Pitman M.D. (Novembro de 2006). Limited Evolutionary Potential. Página visitada em 14 de Agosto de 2012.
- ↑ The Steppingstone Problem: And the Limits of Evolutionary Potential by Sean D. Pitman, M.D. March 2004.
- ↑ Phylogenetic analyses of the constituents of Type III protein secretion systems. J Mol Microbiol Biotechnol. 2 (2), 125-144, 2000.
Ligações externas
Criacionista
- The Bacterial Flagellum por Design and the Universe
- Evolving the Bacterial Flagellum Through Mutation and Cooption por Mike Gene
- The Evolution of the Flagellum: And the Climbing of "Mt. Improbable " por Sean D. Pitman, M.D.
Secular
- [1] Wikipedia - Flagelo
